尼帕病毒(Nipah virus, NiV)是一种高致病性的人畜共患病毒,被世界卫生组织(WHO)列为重点研究的优先病原体。自1998年首次发现以来,该病毒在东南亚和南亚地区引发了多次致命性疫情,病死率高达40%-75%,并表现出人际传播的能力,构成了严重的公共卫生威胁。本文全面综述了尼帕病毒的研究进展,系统分析了其流行病学特征、传播途径、自然宿主生态、临床表现与发病机制。重点阐述了病毒进入、复制和组装的分子生物学机制,包括病毒糖蛋白与宿主受体的相互作用、RNA聚合酶复合物的结构与功能,以及病毒蛋白如何拮抗宿主免疫应答。此外,本文还系统梳理了疫苗和治疗药物的最新研发进展,涵盖了亚单位疫苗、病毒载体疫苗、mRNA疫苗等多种技术路线,以及以中和抗体和瑞德西韦为代表的治疗性药物。最后,本文讨论了当前面临的挑战,如病毒的遗传多样性、散发性暴发的临床试验困境以及新发亨尼帕病毒的威胁,并对未来的研究方向和“同一健康”(One Health)防控策略进行了展望,旨在为全球应对尼帕病毒威胁提供科学参考。 1. 引言尼帕病毒(Nipah virus, NiV)是一种新兴的、由动物传播给人类的高致病性副粘病毒,对其自然宿主果蝠(Pteropus属)通常不致病,但能感染包括人类在内的多种哺乳动物,并引发严重甚至致命的疾病 [1, 2]。该病毒于1998年末至1999年间在马来西亚和新加坡暴发,首次被确认为一种新型病原体。当时,疫情主要在与猪有密切接触的人群中引发了严重的脑炎,最终导至马来西亚报告了超过265例脑炎病例,其中105例死亡 [1, 3]。电子显微镜、血清学和遗传学研究表明,NiV属于副粘病毒科(Paramyxoviridae),与此前在澳大利亚发现的亨德拉病毒(Hendra virus, HeV)亲缘关系最近,两者共同构成了亨尼帕病毒属(Henipavirus) [1]。研究迅速追溯到其自然宿主为当地的果蝠 [4, 3]。 由于尼帕病毒病(Nipah virus disease, NVD)起病急、病死率高,尼帕病毒被归类为生物安全四级(Biosafety Level 4, BSL-4)病原体,其研究和操作必须在最高级别的生物安全防护设施中进行 [5, 6]。鉴于其流行潜力、高致病性以及医疗对策的缺乏,世界卫生组织(WHO)在其“研发蓝图”中将尼帕病毒列为急需加速研究和开发的重点疾病之一 [7, 8, 9]。 自首次发现以来,尼帕病毒已在多个国家和地区引发了数次暴发。最初的马来西亚疫情主要通过受感染的猪作为中间宿主传播给人类 [1, 10]。邻近的新加坡也报告了11例病例,主要发生于曾处理过从马来西亚进口猪的屠宰场工人中 [11]。此后,疫情的地理分布和传播模式发生了显著变化。自2001年起,孟加拉国和印度经历了几乎每年一次的季节性暴发 [12, 13]。在这些地区,病毒的主要传播途径被认为是人类食用了被蝙蝠尿液或唾液污染的生椰枣汁,并且出现了显著的人际传播 [12, 14]。最近,印度喀拉拉邦在2023年再次经历了疫情,临床表现为急性呼吸窘迫综合征和脑炎综合征的混合特征 [15]。 尼帕病毒对全球公共卫生构成了严峻挑战,其核心难点在于其极高的病死率,在不同疫情中可高达40%至75%以上 [14, 13]。此外,病毒具备人际传播的能力,尤其是在孟加拉国的疫情中,约一半的病例是通过人际接触感染的,这增加了其引发更大规模流行的风险 [16]。感染者不仅面临急性呼吸道或神经系统疾病的威胁,部分幸存者还可能出现复发性或迟发性脑炎等长期神经系统后遗症 [17]。当前,全球尚无任何已获批准用于人类的尼帕病毒疫苗或特效治疗药物,这使得预防和控制工作面临巨大压力,相关医疗对策的研发成为全球科研领域的当务之急 [8]。 本综述旨在系统性地总结当前关于尼帕病毒的研究进展。我们将从病毒的分子生物学特征、流行病学与传播动力学、致病机制、宿主免疫应答、临床诊断技术等方面进行深入阐述。同时,本文将重点评述近年来在疫苗研发和治疗策略探索方面取得的突破与面临的挑战,以期为未来有效应对尼帕病毒威胁提供科学参考和方向。 2. 流行病学与传播尼帕病毒不同暴发模式的流行病学与传播特征对比
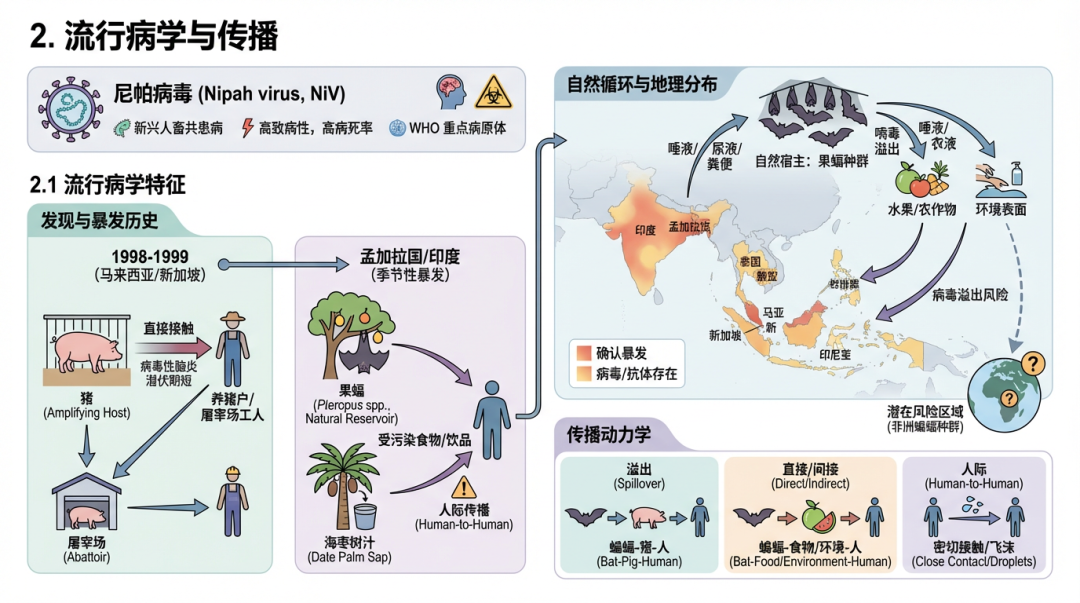
尼帕病毒(Nipah virus, NiV)是一种新兴的、高致病性的人畜共患副粘病毒,因其高病死率和人际传播潜力而被世界卫生组织(WHO)列为重点研究的病原体。本部分将详细阐述尼帕病毒的流行病学特征、自然循环和传播动力学。
2.1 流行病学特征发现与暴发历史:尼帕病毒于1998-1999年在马来西亚首次被确认,当时在与猪有密切接触的养猪户和屠宰场工人中暴发了严重的病毒性脑炎 [18, 1, 3]。流行病学调查显示,患者在发病前两周通常与病猪有过直接接触,提示病毒由猪直接传播给人类,且潜伏期较短 [18]。不久后,新加坡也报告了与从马来西亚进口的猪相关的屠宰场工人感染病例 [10, 28]。 地理分布:尼帕病毒的地理分布主要集中在南亚和东南亚。除了马来西亚和新加坡,孟加拉国和印度也经历了反复的、季节性的暴发 [19, 29]。此外,在柬埔寨、泰国、印度尼西亚和菲律宾等国的蝙蝠种群中也发现了病毒或其抗体的存在 [30, 31, 32, 33]。最近的地球空间模型分析整合了1998年至2021年的人类和动物感染数据,进一步绘制了尼帕病毒的潜在风险区域 [34]。值得注意的是,系统发育分析和血清学证据表明,与亨尼帕病毒相关的病毒也在非洲大陆的蝙蝠种群中传播,并已发生向人类的溢出事件,这极大地扩展了该病毒属的潜在地理范围 [35, 36]。 暴发模式与疾病负担:尼帕病毒暴发呈现出不同的模式。在孟加拉国,暴发几乎每年都在特定季节(12月至次年5月)发生,通常由独立的溢出事件引起 [19, 37]。尼帕病毒感染导至极高的病死率,通常在40%至75%之间,部分暴发中甚至更高 [38, 39, 14]。一项研究发现,在马来西亚暴发期间,脑脊液(CSF)中分离出病毒与高死亡率显著相关,表明中枢神经系统中的高病毒复制可能是导至死亡的重要因素 [40]。此外,由于监测系统主要依赖于医院,许多病例可能未被发现,导至对尼帕病毒真实疾病负担的低估。一项基于医疗求助行为的模型估计,孟加拉国实际发生的暴发数量可能远超官方报告 [41]。幸存者也可能面临长期的神经功能障碍和致残性疲劳等后遗症,增加了疾病的总体负担 [42]。 2.2 自然宿主与病毒循环自然宿主:狐蝠属(Pteropus)的果蝠被确定为尼帕病毒的主要自然宿主 [4, 25, 43]。在马来西亚(P. hypomelanus 和 P. vampyrus)、孟加拉国(P. medius)、印度(P. giganteus)、柬埔寨(P. lylei)等多个国家的多种狐蝠体内都检测到了尼帕病毒的抗体或病毒RNA,证实了其作为储存宿主的广泛性 [25, 26, 27, 44, 30]。蝙蝠对这些高致病性病毒具有独特的耐受性,这可能与其特殊的免疫系统有关 [45]。 病毒在蝙蝠中的动态:尼帕病毒在蝙蝠种群中的动态对其向人类的溢出有直接影响。研究表明,病毒在蝙蝠中的时空动态、血清学动态和遗传学特征共同决定了人类暴发的时间和空间分布 [37]。病毒主要通过蝙蝠的尿液和唾液排出 [31],在人类病例发生地附近的蝙蝠栖息地,可在病例暴露后的长达52天内从蝙蝠尿液中检测到病毒RNA,尽管检测概率随时间迅速下降 [46]。此外,蝙蝠的生理状况(如食物压力)可能会影响其免疫反应和病毒排出。一项研究发现,限制蛋白质摄入会改变蝙蝠的抗体反应,这可能对病毒的维持和传播产生影响 [47]。 病毒进化:基因组测序和系统发育分析揭示了尼帕病毒存在两个主要的遗传谱系:马来西亚株(NiV-M)和孟加拉国株(NiV-B) [20, 21]。这两个谱系在地理和时间上交织在一起,显示了病毒的持续演化和传播 [48]。2018年印度喀拉拉邦暴发的病毒序列与孟加拉国株高度相似,并与当地的Pteropus medius蝙蝠中的病毒序列几乎完全一致,证实了此次暴发的蝙蝠来源 [22]。尽管病毒在不断进化,但一项对受体结合蛋白(RBP)的深入突变扫描研究发现,在自然流行的尼帕病毒株中,能逃逸中和抗体的抗原性突变很少,这为开发广谱抗体疗法和疫苗提供了重要信息 [49]。 2.3 传播动力学尼帕病毒的传播链包括从自然宿主到中间宿主或人类的“溢出”事件,以及随后的人际传播。 人畜共患传播(溢出事件):
人际传播:
3. 临床表现与发病机制尼帕病毒(NiV)关键蛋白/分子及其功能总结
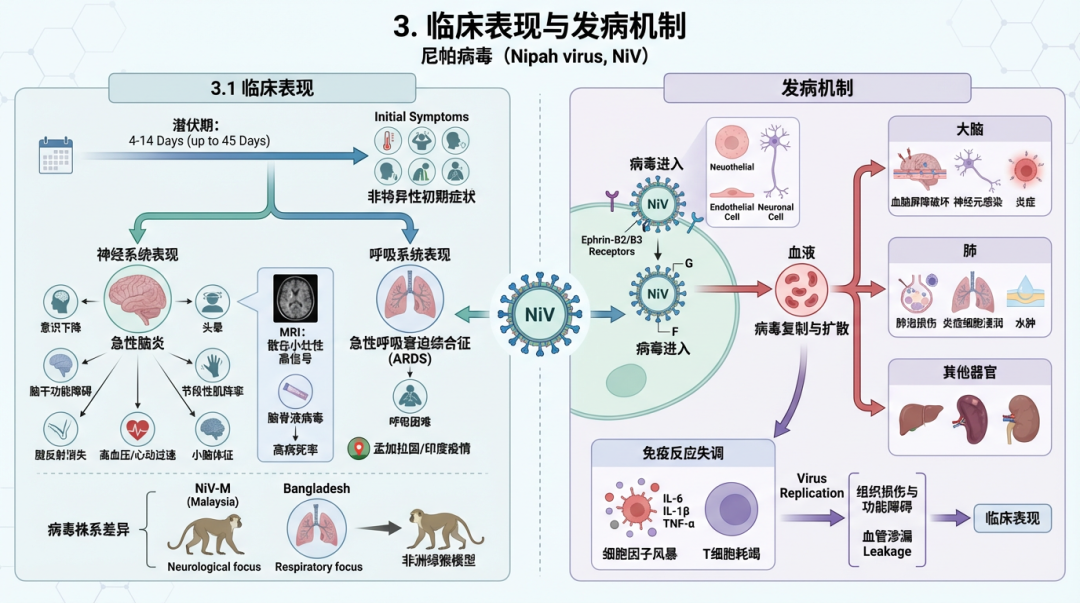
尼帕病毒(Nipah virus, NiV)感染可导至一种严重的、常为致死性的人畜共患病,其临床表现多样,病理过程复杂。本部分将深入探讨NiV感染的临床特征、疾病进程以及潜在的病理生理机制。
3.1 临床表现NiV感染的潜伏期通常为4至14天,但也有长达45天的报道 [18, 83]。疾病初期症状不具特异性,主要表现为发热、头痛、咳嗽、呕吐和呼吸困难等 [14, 18]。 神经系统表现:急性脑炎是NiV感染最严重的临床表现,尤其在马来西亚疫情中最为突出 [18]。患者可迅速出现意识水平下降、头晕、以及显著的脑干功能障碍体征,如节段性肌阵挛、腱反射消失、高血压和心动过速 [18, 84]。小脑体征也较为常见 [84]。脑脊液中检测到病毒与高病死率密切相关 [40]。磁共振成像(MRI)检查显示,急性期患者脑白质、皮层及脑干等部位可见多发、散在的小灶性高信号病变 [85, 84]。值得注意的是,部分无症状的血清阳性者也可能在MRI上出现类似脑部病灶 [86]。 呼吸系统表现:急性呼吸窘迫综合征(ARDS)是另一个主要临床特征,尤其在孟加拉国和印度的疫情中更为常见 [14, 83]。不同病毒株系可能导至不同的临床偏重。在非洲绿猴模型中,与马来西亚株(NiV-M)相比,孟加拉国株(NiV-B)感染导至更高的病死率,并伴有更迅速的呼吸功能衰竭和全身性病毒播散 [80]。 复发与迟发性脑炎:NiV感染的一个独特之处在于其可能导至神经系统后遗症。部分急性脑炎幸存者可在数月至数年后出现复发性脑炎 [17]。此外,初次感染为无症状或非脑炎性疾病的患者也可能出现迟发性脑炎 [17, 87]。对非人灵长类动物幸存者的研究发现,NiV可在脑部持续存在,尤其是在神经元和小胶质细胞中,这为复发和迟发性神经系统疾病提供了可能的病理学基础 [82]。 长期后遗症:幸存者常报告持续性症状,包括致残性疲劳、持续性神经功能障碍(如静态脑病、眼动神经麻痹)等 [42]。 3.2 发病机制NiV的发病机制涉及病毒与宿主相互作用的多个层面,包括高效的细胞入侵、独特的病毒复制与组装策略,以及强大的免疫逃逸能力。 3.2.1 病毒入侵与趋向性NiV具有广泛的宿主嗜性,其关键在于病毒通过其附着糖蛋白(G)与宿主细胞表面的受体结合。已确认的主要受体是Ephrin-B2和Ephrin-B3,这两种蛋白在多种哺乳动物中高度保守,并广泛表达于血管内皮细胞和神经元,这与NiV感染引起的主要病理改变(血管炎和脑炎)部位高度一致 [64, 65, 78]。研究发现,NiV对动脉血管的嗜性高于静脉,这与动脉内皮细胞上更高的Ephrin-B2表达水平相关 [6]。此外,硫酸乙酰肝素(Heparan sulfate)也被发现可作为一种附着因子,增强病毒感染效率 [79]。 病毒的入侵由G蛋白和融合糖蛋白(F)协同完成。G蛋白首先与受体结合,这一过程会诱导G蛋白发生一系列构象变化,包括头部结构域的变构和茎部区域的暴露,最终触发F蛋白发生构象重排,介导病毒包膜与细胞膜的融合 [66, 88]。研究表明,G蛋白四聚体与受体的结合模式是动态的,并且G蛋白介导的Ephrin-B2受体聚集是有效激活F蛋白的必要步骤 [89, 90]。F蛋白在融合前以一种独特的“三聚体六聚体”高级结构存在,这种结构对于稳定其融合前构象和调控膜融合过程至关重要 [91]。病毒感染后,F和G蛋白在细胞表面表达,导至细胞-细胞融合,形成多核巨细胞(合胞体),这是NiV感染的标志性病理特征 [65]。 3.2.2 病毒复制与组装作为一种非节段性负链RNA病毒,NiV的基因组复制和转录由RNA依赖的RNA聚合酶(RdRp)复合体完成。该复合体由大蛋白(L)和磷蛋白(P)组成。近期的多项冷冻电镜(Cryo-EM)研究揭示了NiV L-P复合体的精细结构,阐明了四聚体P蛋白如何与L蛋白的RdRp结构域相互作用,以及RNA模板和新生链在聚合酶活性位点的结合方式,为靶向聚合酶的抗病毒药物(如瑞德西韦)开发提供了结构基础 [67, 92, 93, 94, 95, 68]。 病毒基因组RNA被核蛋白(N)包裹形成核衣壳(RNP)。在这一过程中,P蛋白扮演着分子伴侣的角色,它与游离的N蛋白(N⁰)结合,阻止其自我聚合,并使其保持一种开放的构象,以便随时抓取并包被新合成的病毒RNA [69]。 病毒的组装和出芽主要由基质蛋白(M)驱动。超高分辨率显微镜研究揭示了一种随机组装模型:M蛋白在质膜上聚集形成病毒样颗粒(VLPs),而F和G糖蛋白则以随机方式被包裹进新生的病毒颗粒中,而非由M蛋白主动招募 [70]。M蛋白通过与质膜上的特定脂质(如磷脂酰丝氨酸和PI(4,5)P2)相互作用来驱动病毒组装 [96],并利用皮层肌动蛋白网络来稳定病毒组装位点,促进出芽过程 [97]。此外,M蛋白的核-质穿梭对其功能至关重要,这一过程受到泛素化修饰的精确调控 [98, 99]。病毒的C蛋白也能通过招募ESCRT途径的Tsg101蛋白来促进病毒的有效释放 [77]。 3.2.3 免疫逃逸与宿主相互作用NiV具有强大的免疫抑制能力,这是其高致病性的关键因素。病毒P基因通过RNA编辑和不同的起始密码子,编码P、V、W和C四种蛋白质,其中V和W蛋白是主要的免疫拮抗因子。 在雪貂模型中,V蛋白被证实是决定病毒致病性的主要因素,V蛋白缺陷的病毒毒力显著减弱;而W蛋白则主要调节宿主的炎症反应,W蛋白缺陷的病毒导至病程改变,表现为呼吸道疾病减轻,但晚期神经系统疾病加重 [74]。分子机制研究表明,V蛋白通过与蛋白磷酸酶1(PP1)相互作用,阻止宿主抗病毒感应蛋白MDA5的去磷酸化激活,从而逃避I型干扰素(IFN-I)的产生 [75]。W蛋白则通过与宿主14-3-3蛋白结合,抑制NF-κB信号通路,从而减弱促炎反应 [76]。此外,NiV蛋白还通过一种保守机制拮抗STAT1信号,阻断IFN下游的抗病毒效应 [100]。 除了P基因产物,M蛋白也展现出多重免疫调节功能。它能通过靶向并降解E3泛素连接酶TRIM6来抑制IKKε激酶介导的IFN-I应答 [71]。M蛋白还能劫持宿主细胞核仁蛋白Treacle,抑制核糖体RNA的生物合成 [72],并将m6A甲基转移酶METTL3重定位至细胞质,以增强病毒N蛋白mRNA的稳定性,从而促进病毒复制并抑制宿主免疫 [73]。 在宿主方面,对人类幸存者的研究显示,急性感染期存在显著的活化CD8+ T细胞增多,同时伴有病毒特异性IgM和IgG抗体的产生,表明细胞免疫和体液免疫均参与了病毒清除过程 [101]。然而,病毒感染导至的血管内皮损伤和合胞体形成是主要的病理特征 [18]。最近的研究发现,NiV诱导的合胞体形成会导至线粒体膜电位丧失和线粒体DNA泄漏至细胞质,进而通过cGAS和IFI16激活STING信号通路,诱导IFN-β和IL-6的产生,这揭示了病毒诱导的细胞融合与宿主先天免疫应答之间的新联系 [81]。多组学分析也表明,NiV糖蛋白能够调节线粒体相关蛋白,并改变细胞的糖、脂质和氨基酸代谢,以支持病毒复制所需的能量供应 [102]。这些复杂的相互作用共同决定了NiV感染的临床进程和最终结局。 4. 病毒分子生物学尼帕病毒(Nipah virus, NiV)属于副粘病毒科(Paramyxoviridae)亨尼帕病毒属(Henipavirus),是一种人畜共患的、有包膜的负链单股RNA病毒。其基因组长约18.2 kb,编码至少六种结构蛋白:核蛋白(N)、磷蛋白(P)、基质蛋白(M)、融合蛋白(F)、附着糖蛋白(G)和大的聚合酶蛋白(L),以及通过P基因的RNA编辑产生的V、W和C蛋白 [103]。这些蛋白质协同作用,完成了病毒从入侵宿主细胞到复制和释放的整个生命周期。
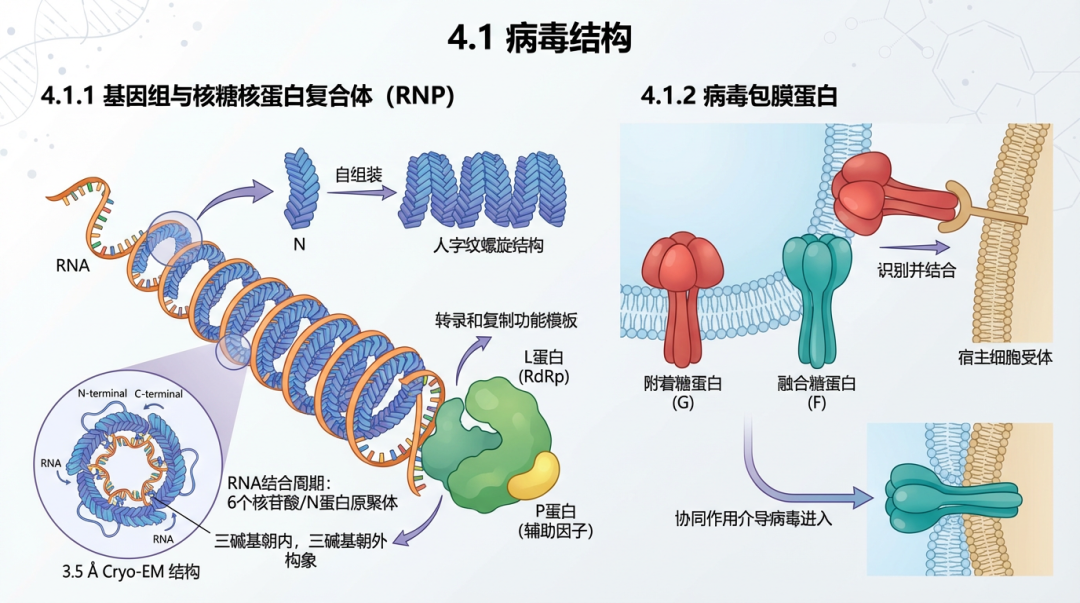
4.1 病毒结构尼帕病毒结构蛋白及其功能总结
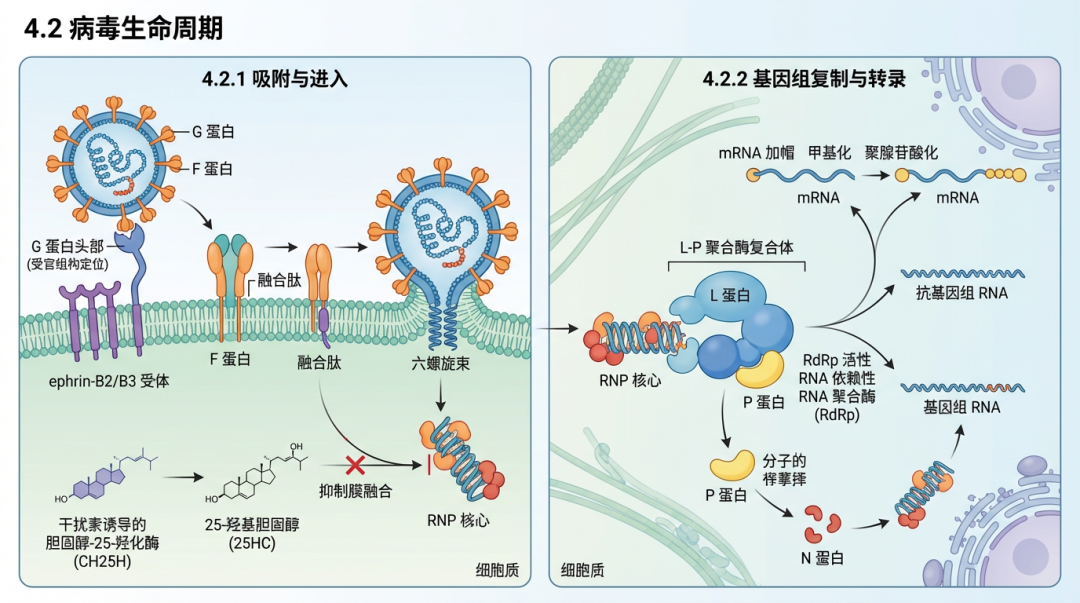
4.1.1 基因组与核糖核蛋白复合体 (RNP)尼帕病毒的基因组RNA被核蛋白(N蛋白)紧密包裹,形成一个具有螺旋对称性的核衣壳,即核糖核蛋白复合体(RNP)。这个RNP复合体是病毒转录和复制的功能模板。N蛋白在体外表达时能够自组装成类似人字纹的螺旋结构 [104, 105]。最近的冷冻电镜(cryo-EM)研究以3.5 Å的分辨率解析了尼帕病毒核衣壳的结构,揭示了其N端和C端区域在稳定螺旋组装中的关键作用。该结构显示,RNA分子以每个N蛋白原聚体结合六个核苷酸的周期性方式缠绕在蛋白复合体上,并呈现出“三碱基朝内,三碱基朝外”的独特构象,这种构象通过蛋白的可塑性实现了对RNA的非序列特异性识别 [106]。这种结构与其它副粘病毒如副流感病毒5(PIV5)的N-RNA复合体结构模型具有相似性 [115]。RNP复合体还包含作为病毒RNA依赖性RNA聚合酶(RdRp)的L蛋白和作为其辅助因子的P蛋白。 4.1.2 病毒包膜蛋白尼帕病毒的包膜上有两种关键的跨膜糖蛋白——附着糖蛋白(G)和融合糖蛋白(F),它们协同作用介导病毒进入宿主细胞。 附着糖蛋白(G蛋白):G蛋白负责识别并结合宿主细胞表面的受体,主要是埃夫林B2(ephrin-B2)和埃夫林B3(ephrin-B3)[107]。G蛋白以同源四聚体的形式存在于病毒表面。冷冻电镜结构分析显示,NiV G蛋白的四聚体胞外域结构复杂,其受体结合头区是引发宿主体内多克隆抗体反应的主要免疫优势区域 [108]。晶体结构研究首次揭示了副粘病毒附着蛋白与细胞蛋白受体相互作用的细节,显示ephrin-B3的一个突出环状结构与NiV-G蛋白表面形成广泛的蛋白-蛋白相互作用界面 [107]。这一结合模式在不同亨尼帕病毒株之间高度保守,即使是遗传上差异较大的非洲亨尼帕病毒,其G蛋白也能以相似的方式识别并结合人类ephrinB2受体 [116, 117]。G蛋白是中和抗体的主要靶点,许多高效价的中和抗体通过阻断其受体结合界面来发挥作用 [109, 118]。然而,一些新发现的亨尼帕病毒,如琅琊病毒(Langya virus, LayV),其G蛋白结构独特,呈蘑菇状,且不与ephrin B2/B3结合,表明亨尼帕病毒属内存在受体使用的多样性 [119, 120]。 融合糖蛋白(F蛋白):F蛋白属于I类病毒融合蛋白,以同源三聚体的形式存在。它在合成后被宿主蛋白酶切割成F1和F2两个亚基,并保持在一种亚稳态的融合前构象(prefusion conformation)。当G蛋白与受体结合后,F蛋白被触发,发生剧烈的构象重排,转变为稳定的融合后构象(postfusion conformation)。在此过程中,F蛋白的疏水性融合肽插入宿主细胞膜,最终形成一个稳定的六螺旋束(6-HB)结构,将病毒膜和细胞膜拉近并融合 [110]。F蛋白的融合前构象是诱导强效中和抗体反应的关键,这些抗体通常识别其顶端的四级表位,通过锁定其融合前构象来阻止膜融合过程 [111, 121, 112]。冷冻电镜技术已成功解析了亨德拉病毒F蛋白的融合前构象晶体结构 [113],以及多种针对NiV F蛋白不同脆弱表位的中和抗体复合物结构,极大地扩展了我们对F蛋白抗原性的认识 [122]。超高分辨率显微镜研究还发现,NiV-F蛋白在细胞膜上并非均匀分布,而是形成独特的纳米级簇,这种聚集状态有利于促进膜融合的触发 [123]。 基质蛋白(M蛋白):M蛋白位于病毒包膜内侧,连接着RNP和包膜糖蛋白的胞内尾部,是病毒组装和出芽过程的组织者。研究表明,M蛋白的核质穿梭对其功能至关重要,这一过程在多种副粘病毒中是保守的 [114]。 4.2 病毒生命周期病毒生命周期中的关键蛋白/分子及其功能
4.2.1 吸附与进入病毒的生命周期始于G蛋白与宿主细胞表面的ephrin-B2/B3受体结合。这一结合事件会诱导G蛋白发生构象变化,特别是暴露其茎部区域,进而触发相邻的F蛋白 [125]。在副粘病毒中,附着蛋白和融合蛋白通常形成一个功能复合体,其中附着蛋白的头部负责识别受体,而其与F蛋白的相互作用则调控融合的启动 [124]。F蛋白被激活后,其融合肽插入靶细胞膜,随后F蛋白发生不可逆的构象重排,形成六螺旋束,最终导至病毒包膜与细胞膜融合,将病毒的RNP核心释放到细胞质中。这一过程可被多种宿主因子或小分子化合物抑制,例如,干扰素诱导的胆固醇-25-羟化酶(CH25H)产生的25-羟基胆固醇(25HC)能够广泛抑制包括尼帕病毒在内的多种包膜病毒通过阻断膜融合进入细胞 [126]。 4.2.2 基因组复制与转录进入细胞质后,病毒的RNP作为模板,在L-P聚合酶复合体的催化下进行RNA的合成。L蛋白是一个多功能酶,具有RNA依赖性RNA聚合酶(RdRp)、mRNA加帽及甲基化、以及聚腺苷酸化等多种活性。P蛋白作为L蛋白的辅助因子,同时还作为N蛋白的分子伴侣,将新合成的N蛋白维持在无RNA结合的单体状态(N0),以供基因组复制时包裹新生RNA。 近期多项冷冻电镜研究成功解析了尼帕病毒L-P聚合酶复合体的近原子分辨率结构 [128, 129, 130]。这些结构揭示了L蛋白的RdRp和多聚核糖核苷酸转移酶(PRNTase)等核心功能域的组织方式,以及P蛋白四聚体与L蛋白的相互作用模式。这些研究为理解负链RNA病毒保守的转录和复制机制提供了重要见解,例如mRNA的共转录成熟通路,包括加帽、甲基化和加A尾等步骤,均由L蛋白独立完成 [127]。此外,这些高分辨率结构也为抗病毒药物的研发提供了精确靶点。例如,针对麻疹病毒聚合酶的非核苷抑制剂ERDRP-0519,被预测并证实对尼帕病毒聚合酶同样有效,其结构研究揭示了该抑制剂通过变构效应将聚合酶锁定在无活性的构象 [131]。病毒RNA的合成发生在细胞质内的特定区域,称为包涵体(IBs)。尼帕病毒感染会形成两种不同类型的包涵体:一种位于核周区域(IBperi),另一种位于质膜下(IBPM),后者含有M蛋白,可能与病毒的组装位点有关 [132]。 4.2.3 组装、出芽与免疫逃逸新合成的病毒基因组RNP、M蛋白和包膜糖蛋白(F和G)在细胞质膜的特定区域聚集。M蛋白在其中起着关键的组织作用,它与RNP和糖蛋白的胞内域相互作用,驱动病毒粒子的组装和出芽。 除了结构蛋白,P基因还通过RNA编辑和替代开放阅读框翻译产生V、W和C等非结构蛋白。这些蛋白是重要的毒力因子,主要功能是拮抗宿主的先天性免疫应答。例如,W蛋白能够进入细胞核,其核输入过程对特定的输入蛋白α3具有高度特异性 [133],且其本身为内在无序蛋白 [134]。此外,多种副粘病毒的V蛋白被发现可以与宿主蛋白TRIM28相互作用,这可能是病毒逃逸宿主防御的一种策略 [135]。这些免疫逃逸机制使得尼帕病毒能够在宿主体内有效复制和传播。 5. 预防与治疗对策鉴于尼帕病毒(NiV)的高致病性和大流行潜力,开发有效的预防和治疗手段是全球公共卫生的当务之急。目前尚无获批用于人体的尼帕病毒疫苗或特效治疗药物,因此相关研究主要集中在疫苗开发、抗病毒药物筛选和公共卫生干预三个方面。
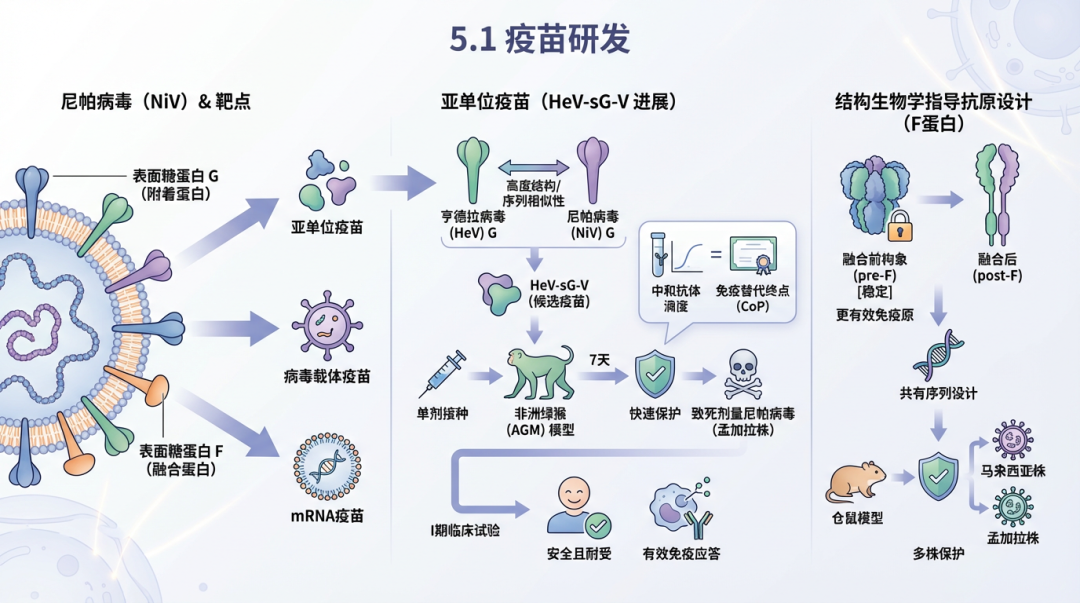
5.1 疫苗研发尼帕病毒疫苗研发策略总结
尼帕病毒的疫苗研发主要以其表面糖蛋白G(附着蛋白)和F(融合蛋白)为靶点,涵盖了亚单位疫苗、病毒载体疫苗和mRNA疫苗等多种技术路线。 亚单位疫苗是研究最为广泛的策略之一。其中,基于亨德拉病毒可溶性G糖蛋白(HeV-sG)的亚单位疫苗(HeV-sG-V)是进展最快的候选疫苗之一。由于亨德拉病毒与尼帕病毒在G蛋白上具有高度的结构和序列相似性,HeV-sG-V被证实能诱导针对尼帕病毒的交叉保护性免疫 [136]。在非洲绿猴(AGM)模型中,单剂接种HeV-sG-V即可在7天内提供对致死剂量尼帕病毒(孟加拉株)攻击的快速保护 [137],并且已确定了与保护相关的中和抗体滴度,为疫苗的审批提供了关键的免疫替代终点(correlate of protection, CoP)[138]。该疫苗已进入I期临床试验,结果显示其在健康成人中具有良好的安全性和耐受性,并能有效诱导针对尼帕病毒的免疫应答 [136]。 为了增强免疫原性和稳定性,研究人员利用结构生物学指导抗原设计。例如,通过稳定F蛋白的融合前构象(pre-F),可以开发出更有效的免疫原 [139, 140]。基于F蛋白共有序列设计的疫苗在仓鼠模型中也显示出对不同尼帕病毒株(马来西亚株和孟加拉株)的保护效果 [141]。纳米颗粒技术是提升亚单位疫苗效果的另一重要方向。将G蛋白头域或通过内含子剪接技术将G蛋白展示在铁蛋白纳米颗粒(ferritin nanoparticles, FeNP)表面,构建的疫苗(NiV G-ferritin)在仓鼠模型中诱导了更快、更广谱、更高滴度的中和抗体,并提供了完全保护 [142, 143, 144]。此外,利用无细胞蛋白合成系统将F和G蛋白共翻译折叠到纳米级脂质体中,形成蛋白脂质体疫苗,为快速生产提供了一条新途径 [145]。 病毒载体疫苗也取得了显著进展。基于水疱性口炎病毒(VSV)的重组疫苗是其中的佼佼者。表达尼帕病毒G蛋白的rVSV疫苗(VSV-NiVG)在非人灵长类(NHP)模型中显示出强大的保护效力,能完全抵御同源和异源病毒株的攻击 [146, 147]。该疫苗的一大优势是能快速起效,在NHP模型中,攻击前7天甚至3天接种仍能提供完全或部分保护 [58]。同样,表达亨德拉病毒G蛋白的单循环rVSV疫苗也能在仓鼠模型中提供对亨德拉和尼帕病毒的双重保护 [148]。另一类是腺病毒载体疫苗,如黑猩猩腺病毒载体疫苗(ChAdOx1 NiV, AdC68-F/G)。ChAdOx1 NiV在AGM模型中提供了对致死攻击的完全保护 [149],而AdC68载体疫苗通过鼻内给药可在仓鼠中快速诱导保护性免疫,并能在小鼠中诱导长达68周的持久免疫力 [150, 151]。此外,J副粘病毒也被探索作为一种新型疫苗载体平台 [152]。 mRNA疫苗作为一种新兴技术平台,在尼帕病毒疫苗研发中也展现出巨大潜力。基于脂质纳米颗粒(LNP)递送的编码F蛋白、G蛋白或两者的mRNA疫苗,在小鼠和仓鼠模型中均诱导了强大而持久的免疫反应,并能完全保护仓鼠免受致死剂量的病毒攻击,且对马来西亚和孟加拉两种主要毒株均有效 [153]。在猪模型中的研究也证实,mRNA疫苗能诱导高效价的中和抗体,这对于阻断作为中间宿主的猪的传播链具有“同一健康”(One Health)的重要意义 [154, 155]。 此外,一些创新疫苗策略也在探索中。例如,一种通过靶向抗原呈递细胞表面CD40受体来递送尼帕病毒抗原的疫苗,在NHP模型中诱导了包括IgA在内的全面免疫应答,并实现了针对尼帕病毒-孟加la株的清除性免疫(sterilizing immunity)[156]。基于病毒复制子颗粒(replicon particle)的黏膜疫苗(NiVΔF)也显示出极快的保护效果 [63]。同时,免疫信息学和机器学习等计算方法正被用于预测和设计新型多表位疫苗 [157, 158]。 5.2 治疗性药物尼帕病毒在研治疗性药物总结
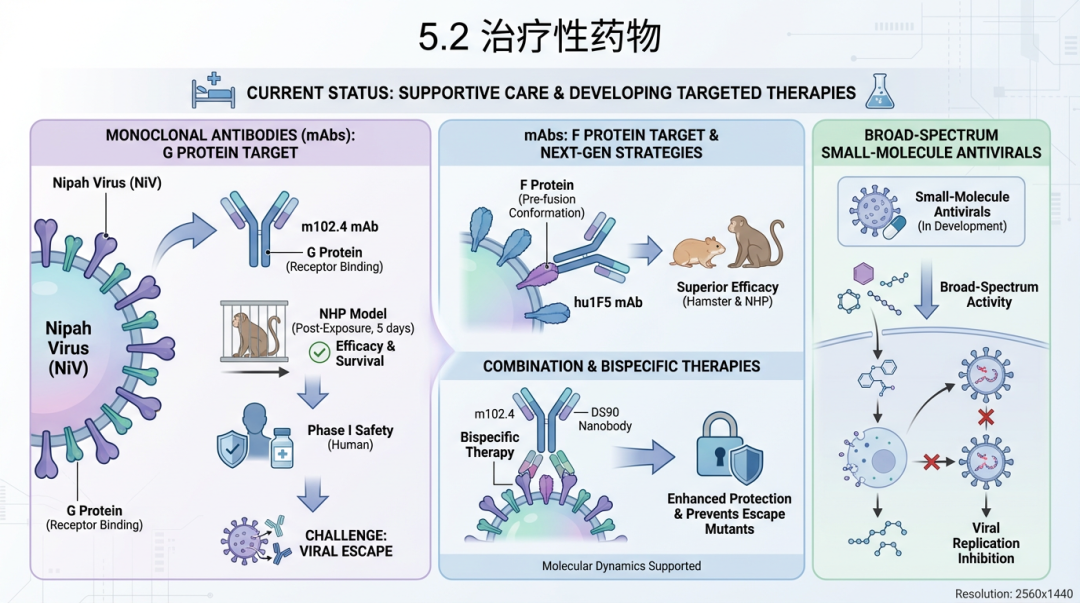
目前,尼帕病毒感染的治疗仍以支持性护理为主,但多种针对性治疗药物的研发正在积极进行中,主要包括单克隆抗体和广谱小分子抗病毒药物。 单克隆抗体(mAbs)是目前最有希望的治疗手段。人源单克隆抗体m102.4是研究最深入的候选药物,它靶向病毒G蛋白的受体结合域,对尼帕病毒和亨德拉病毒均有高效的中和活性。在NHP模型中,即使在病毒暴露后5天(已出现临床症状)开始给予m102.4治疗,仍能显著提高存活率 [159]。一项针对健康成人的I期临床试验表明,m102.4具有良好的安全性和耐受性,为其临床应用铺平了道路 [160]。然而,病毒的免疫逃逸是单抗治疗面临的挑战。 针对F蛋白的单抗是另一个重要的研发方向。研究发现,靶向F蛋白融合前构象的单抗hu1F5在仓鼠和NHP模型中均显示出优于m102.4的治疗效果,表明F蛋白是更优的治疗靶点 [161]。为了应对病毒逃逸,研究人员正在开发组合疗法和新一代抗体。一项研究开发了一种双特异性疗法,将靶向F蛋白的纳米抗体(DS90)与m102.4结合,该疗法在动物模型中不仅提供了更优的保护,还有效地阻止了病毒逃逸突变株的出现 [162]。分子动力学模拟也从理论上证实了不同抗体组合的协同作用机制 [168]。这些研究表明,被动免疫疗法是极具潜力的暴露后预防和治疗策略 [169]。
小分子抗病毒药物的开发也取得了重要进展。广谱抗病毒药物瑞德西韦(Remdesivir, GS-5734)在AGM模型中被证实能有效保护动物免受尼帕病毒的致死性攻击,是主要的候选药物之一 [68]。另一种口服核糖核苷类似物4'-氟尿苷(4'-Fluorouridine, EIDD-2749)在体外对尼帕病毒有抑制活性,并在其他出血热病毒的动物模型中显示出高效力 [163]。利巴韦林(Ribavirin)曾在马来西亚疫情期间的一项开放标签试验中使用,结果显示与死亡率降低相关,但该研究的设计存在局限性,其确切疗效仍需更严格的临床试验证实 [38]。 此外,病毒融合抑制剂是另一类有前景的小分子药物。靶向F蛋白七肽重复区(HR)的抑制剂能够阻断病毒膜融合。例如,胆固醇标记的HRC衍生肽在体内显示出抗病毒活性 [164],而一种新型的去PEG化脂肽在体外实验中表现出皮摩尔(pM)级别的超强抑制活性 [165]。 其他创新疗法也在探索之中。例如,缺陷性干扰颗粒(Defective Interfering Particles, DIPs)是一种利用天然存在的、基因组不完整的病毒颗粒来干扰标准病毒复制的新型生物疗法。在仓鼠模型中,预防性给予尼帕病毒来源的DIPs可显著降低致死率,但暴露后治疗无效,提示其可能更适用于预防而非治疗 [166, 167]。 5.3 公共卫生防控措施尼帕病毒公共卫生防控措施总结
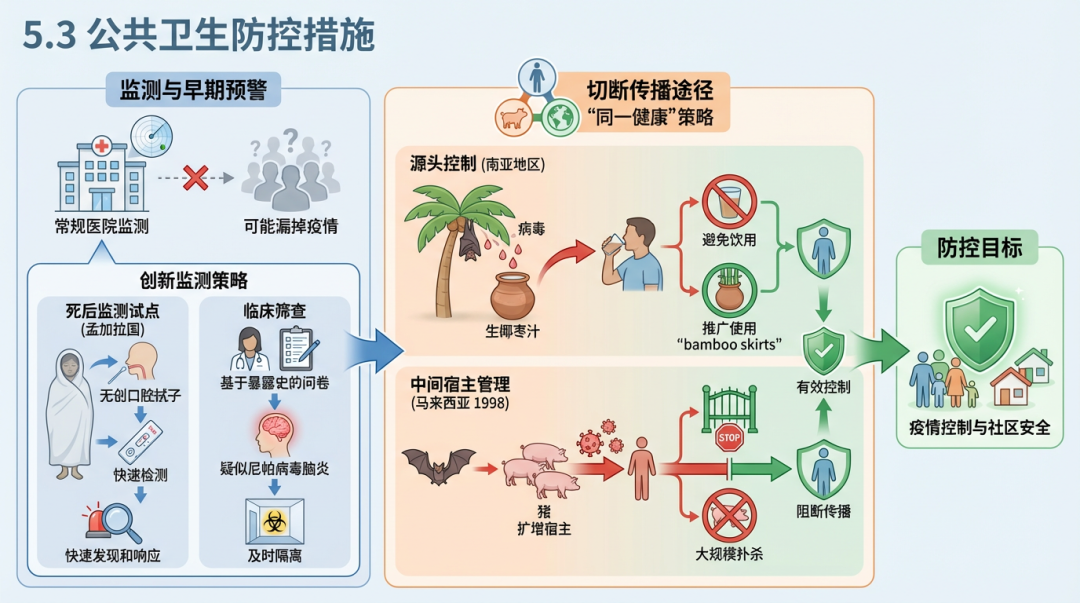
在缺乏有效疫苗和药物的情况下,强有力的公共卫生措施是控制尼帕病毒疫情的关键。这些措施包括加强监测、切断传播途径和管理社会心理影响。 监测与早期预警是防控的第一道防线。研究表明,仅依赖医院的常规监测可能会漏掉大量疫情 [41]。因此,创新的监测策略至关重要。例如,在孟加拉国开展的死后监测(post-mortem surveillance)试点项目,通过对死者进行无创口腔拭子采样和快速检测,成功实现了对疫情的快速发现和响应 [170]。在临床环境中,使用基于暴露史的筛查问卷,可以帮助医护人员更快地识别疑似尼帕病毒脑炎患者,从而及时采取隔离措施 [171]。
切断传播途径是控制疫情的核心。这需要采取“同一健康”的综合策略。
社会心理支持也是公共卫生应对中不可或缺的一环。尼帕病毒等高致死率传染病暴发往往会给社区带来严重的社会污名化。为应对这一问题,研究人员已开发并验证了可用于尼帕病毒等疫情的“新发和流行性传染病(RAPID)污名量表”,以实时监测和干预污名问题 [172]。此外,疫情给幸存者、家属和社区居民带来的创伤后应激障碍(PTSD)等心理健康问题,也需要得到充分的关注和支持 [173]。 6. 挑战与未来展望尼帕病毒研究的挑战与未来展望总结
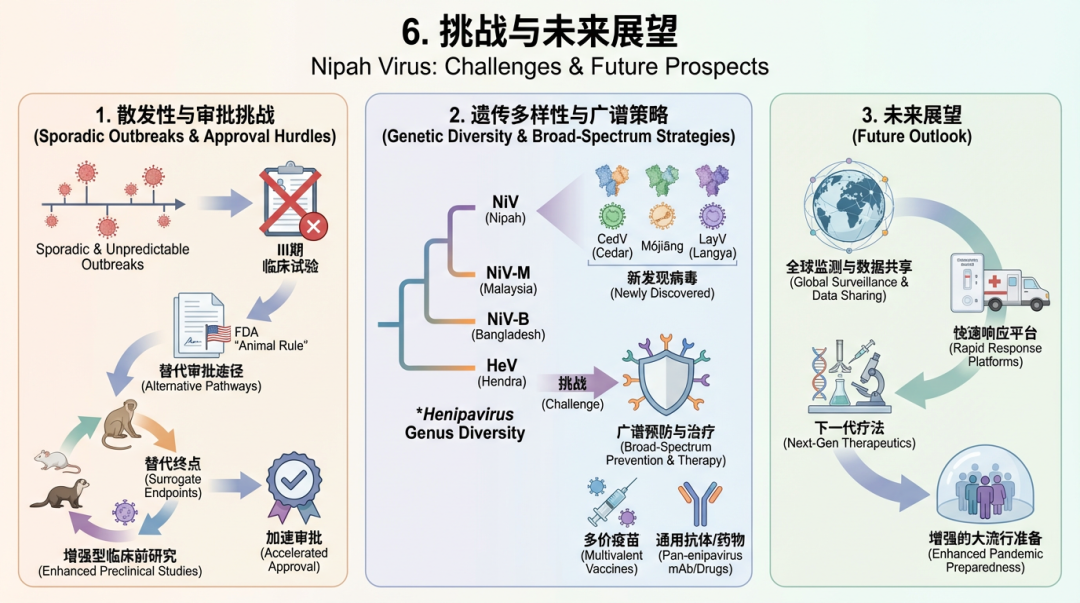
尼帕病毒(Nipah virus, NiV)作为一种高致病性、具有大流行潜力的新发病原体,其研究和防控工作在取得进展的同时,也面临着诸多严峻挑战。
首先,疫情的散发性和不可预测性是当前面临的核心难题之一。尼帕病毒暴发通常规模小、时间短且地理位置分散,这使得开展传统的III期临床有效性试验变得极为困难 [174]。因此,为疫苗和疗法寻找替代的审批途径,例如美国食品药品监督管理局(FDA)的“动物规则”(Animal Rule),成为必然选择,而这反过来也对临床前研究和替代终点的确立提出了更高要求 [138]。 其次,亨尼帕病毒属(Henipavirus)内部巨大的遗传多样性,以及新病毒的不断发现,对广谱预防和治疗策略构成了重大挑战。不仅尼帕病毒本身存在不同的遗传谱系(如马来西亚株和孟加拉国株),近年来在中国、澳大利亚等地陆续发现了与尼帕病毒和亨德拉病毒(Hendra virus, HeV)亲缘关系较远的亨尼帕病毒,如雪松病毒(Cedar virus, CedV)、莫江病毒(Mòjiāng virus)和琅琊病毒(Langya virus, LayV)[175, 176, 188]。结构和抗原性研究表明,这些新发现病毒的表面糖蛋白与经典的尼帕病毒存在显著差异 [177, 119, 120, 189]。例如,琅琊病毒的F蛋白抗原性与尼帕病毒F蛋白截然不同 [177],而雪松病毒则能够利用更多种类的Ephrin受体进行入侵 [178]。这种多样性意味着针对特定尼帕病毒株开发的疫苗或单克隆抗体可能无法提供对其他亨尼帕病毒的有效保护,因此开发广谱性防治手段迫在眉睫 [190, 191]。 最后,我们对尼帕病毒持续性感染和复发性脑炎的发病机制仍知之甚少。临床研究早已发现,部分幸存者在急性感染后数月甚至数年会出现复发性或迟发性脑炎 [17]。非人灵长类动物模型也证实,尼帕病毒能够在急性期后持续存在于中枢神经系统,尤其是在神经元中 [82]。然而,病毒潜伏、持续复制和再次激活的具体分子机制尚不明确。借鉴相关副粘病毒如麻疹病毒的研究,病毒F蛋白发生突变导至其呈现“高融合性”表型,可能是病毒在缺乏经典受体的神经元中高效传播和导至亚急性硬化性全脑炎(SSPE)的关键,这一机制是否同样适用于尼帕病毒的神经系统后遗症,值得深入探究 [192]。 面对上述挑战,未来的研究应聚焦于以下几个方向: 第一,开发广谱抗亨尼帕病毒的疫苗和疗法。未来的研发策略应着眼于病毒的保守靶点。例如,基于结构设计的、能稳定在融合前构象的F蛋白,或包含多种病毒保守表位的嵌合抗原,已被证明能诱导针对不同亨尼帕病毒株(包括尼帕病毒和亨德拉病毒)的交叉中和抗体 [121, 139, 193]。通过理性设计改造病毒糖蛋白,重建或暴露交叉反应性抗原表位,是实现广谱保护的有效途径 [194, 195]。近期发现的能同时中和尼帕病毒和琅琊病毒的抗体,更揭示了开发真正广谱疗法的可能性 [179]。此外,一些基于mRNA、重组蛋白和病毒载体的新型疫苗平台也显示出良好的交叉保护潜力 [196, 197]。 第二,建立可靠的疫苗保护性免疫相关指标(correlates of protection, CoPs)。鉴于临床有效性试验的困难,确定能够预测疫苗保护效果的免疫学指标对于加速疫苗审批至关重要。近期在非人灵长类动物模型中的研究取得了重要突破,成功将特定的中和抗体滴度与动物在致死剂量病毒攻击下的存活率关联起来,为尼帕病毒疫苗的评估和审批提供了明确的科学依据和基准 [138, 180]。 第三,利用高通量筛选和人工智能等新技术加速抗病毒药物的发现。传统药物研发周期长,难以应对突发疫情。构建新型高通量筛选平台,如基于转录和复制能力的病毒样颗粒(trVLPs)系统或无细胞的聚合酶活性检测系统,能够在生物安全二级(BSL-2)条件下快速筛选大规模化合物库,寻找潜在的抗病毒药物 [129, 181]。同时,人工智能、机器学习和知识库构建等计算方法可用于药物的虚拟筛选、先导化合物的优化,甚至预测病毒的进化和逃逸突变,从而指导开发更持久有效的疗法 [182, 183, 198, 199]。 第四,加强对蝙蝠的病毒生态学和免疫学研究,以预测和预防病毒溢出。作为亨尼帕病毒的天然宿主,对蝙蝠的研究是从源头防控的关键。这需要通过宏转录组学等手段,在全球范围内加强对蝙蝠种群中亨尼帕病毒遗传多样性和地理分布的监测 [184, 37, 200]。深入研究影响病毒在蝙蝠种群中传播的生态驱动因素,如宿主饮食、栖息地环境、季节变化以及与人类活动的交集,有助于建立溢出风险的预测模型 [54, 184]。此外,揭示蝙蝠独特的免疫系统如何与病毒共存,例如其特殊的干扰素反应 [185] 以及营养胁迫等因素如何影响其免疫状态和病毒排毒,将为预测和预防病毒从动物到人的跨种传播提供至关重要的线索 [47]。 综上所述,应对尼帕病毒及其他新发亨尼帕病毒的威胁,必须采取综合性的策略。人类、动物和环境健康的相互关联性凸显了“同一健康”(One Health)理念的核心地位。这不仅要求生物医学领域的持续投入,还需关注土地利用变化、集约化农业生产模式和气候变化等增加人与野生动物接触的宏观生态因素 [186, 187]。持续的国际合作,在监测、科研和资源共享等方面协同努力,是有效应对未来疫情、保障全球公共卫生安全的基石。 7. 结论作为一种高致病性新兴人畜共患病毒,尼帕病毒(NiV)因其高病死率、广泛的宿主范围和人际传播能力,持续对全球公共卫生构成严重威胁,并被世界卫生组织列为需要优先研发对策的重点病原体 [1, 201]。在过去二十余年中,全球科研界在应对尼帕病毒挑战方面取得了显著进展。 在流行病学和病毒溯源方面,持续的监测极大地扩展了我们对尼帕病毒地理分布、宿主范围和遗传多样性的认识。在菲律宾等地的蝙蝠种群中开展的血清学调查,不仅证实了狐蝠作为其主要自然宿主的地位,还揭示了年龄和季节等因素对病毒血清阳性率的影响 [202]。同时,在比利时等非传统疫区的哺乳动物中发现新型副粘病毒,凸显了在全球范围内开展广泛病毒学监测的重要性 [203]。对不同地区(如印度2018年分离株)的病毒株进行基因组和致病性分析,也加深了我们对病毒演化和毒力差异的理解 [204]。在分子生物学和发病机制研究领域,近期的结构生物学研究取得了突破性进展,特别是利用冷冻电镜技术解析了尼帕病毒聚合酶(L-P)复合物在不同状态下的高分辨率结构,揭示了其在RNA合成过程中的动态重塑机制,为开发靶向聚合酶的抑制剂提供了精确的结构基础 [205, 92, 67]。对感染幸存者的免疫学研究同样成果斐然。研究发现,即使在感染25年后,马来西亚疫情的幸存者体内仍能检测到针对融合蛋白(F)和附着蛋白(G)的持久性抗体及记忆B细胞 [206]。对2023年印度喀拉拉邦疫情幸存者的免疫分型分析,则揭示了持续性淋巴细胞减少等独特的免疫特征,为理解保护性免疫应答和病毒致病机理提供了关键线索 [207, 208]。 在疫苗和治疗药物的研发方面,一系列令人鼓舞的成果为尼帕病毒的防治带来了希望。基于亨德拉病毒可溶性G蛋白(HeV-sG-V)的亚单位疫苗已完成I期临床试验,显示出良好的安全性和免疫原性 [182]。重组水疱性口炎病毒(rVSV)载体疫苗平台也表现出巨大潜力,不仅在非人灵长类动物模型中证实了其能够快速诱导并提供持久的保护效果,而且其安全性评估也取得了积极进展 [58, 209]。此外,研究人员正在积极探索多种创新疫苗策略,包括mRNA疫苗、多价病毒样颗粒疫苗、分子钳亚单位疫苗以及利用动态共价水凝胶等新型佐剂增强免疫应答的单剂疫苗等 [210, 211, 212, 213, 214, 215]。系统性地评估这些不同技术平台(如蛋白亚单位、病毒载体和RNA疫苗)的效力,对于优化未来的疫苗开发至关重要 [216]。在治疗方面,人源单克隆抗体m102.4和小分子药物瑞德西韦(Remdesivir)在动物模型中均显示出显著的保护效果,其中m102.4已在同情用药中得到应用 [217, 159, 68]。同时,基于病毒蛋白结构信息的计算机辅助药物筛选(如发现潜在的喹诺酮类衍生物抑制剂)和利用尼帕病毒蛋白改造的靶向递送系统等前沿研究,也为开发新型疗法开辟了新途径 [218, 219]。然而,相关副粘病毒研究提示,病毒可能通过突变逃避中和抗体,这强调了开发广谱或联合疗法的必要性 [220]。 综上所述,尽管在基础研究和医学对策开发方面取得了长足进步,尼帕病毒作为一种高危新兴病原体的威胁依然存在。未来的挑战要求我们必须采取综合性的“同一健康”(One Health)策略,整合对野生动物、家畜和人类的监测 [221]。这需要全球社会加大投入,强化跨境监测网络,利用新型诊断工具(如Split NanoLuc生物传感器)实现早期预警和精准溯源,并注意与其它病原(如蝙蝠呼肠孤病毒)的鉴别诊断 [222, 223]。同时,必须持续推动基础研究,深入揭示病毒复制、免疫逃逸和跨种传播的分子机制。更重要的是,要加速转化医学的进程,将实验室中的候选疫苗和药物高效地推向临床,并建立合理的储备和分配模型以应对潜在的暴发需求 [224]。通过这些协同努力,我们不仅能更有效地应对尼帕病毒的挑战,也将为抵御未来其他新发和再发传染病奠定坚实的基础。 Supplementary Studies / Further Reading补充研究/延伸阅读除了上文已详细讨论的核心研究,近年来还有大量补充性研究从病毒生态学、宿主-病毒相互作用、新型诊疗策略及结构生物学等多个维度,进一步加深了我们对亨尼帕病毒(尤其是尼帕病毒)的理解。 病毒生态学、传播动力学与风险评估对尼帕病毒(NiV)自然宿主及其溢出机制的深入探索对于预防至关重要。在柬埔寨的一项研究中,尽管在蝙蝠尿液中检测到了NiV RNA,并发现了可能导至病毒传播的人类活动(如食用被蝙蝠污染的水果和生产棕榈汁),但在418名潜在暴露人群中并未发现血清阳性者,这揭示了病毒溢出事件的复杂性和偶然性 [225]。相比之下,孟加拉国的研究明确指出,饮用被蝙蝠污染的生海枣汁是主要的传播途径 [226]。除了直接从蝙蝠溢出,家养动物的潜在作用也受到关注。一项研究在孟加拉国人类疫情发生地的牛、狗和猫体内检测到NiV抗体,提示这些动物可能在病毒传播链中扮演一定角色 [227]。 为了更精准地识别和预测风险,研究人员开始采用创新的方法。例如,利用系统发育因子分解(phylofactorization)这一机器学习算法,可以对现有监测数据进行分析,以识别最有可能携带亨尼帕病毒和丝状病毒的蝙蝠分支,从而指导未来的监测工作 [228]。同时,有模型研究提出,通过植树造林和农林复合经营等环境管理措施,可能有助于减少蝙蝠与人类的接触,从而降低蝙蝠源性病原体(如NiV)的传播风险 [229]。从更宏观的视角看,粮食系统与大流行病之间的联系也日益受到重视,不安全的农业实践和食品处理方式被认为是促进人畜共患病(包括NiV)出现和传播的重要因素 [230]。此外,一项关于WHO蓝图优先疾病暴发预测的范围综述指出,尽管预测模型日益复杂,但对NiV等疾病的准确预测仍面临巨大挑战 [231]。 病毒本身的遗传多样性和致病性也得到了持续关注。对柬埔寨莱氏狐蝠(Pteropus lylei)分离的NiV毒株(CSUR381)的深入分析证实,该毒株属于马来西亚基因型,并在仓鼠模型中表现出高致病性,表明在自然界蝙蝠种群中循环的病毒株具有直接对人类构成威胁的潜力 [232]。对孟加拉国2008-2010年间暴发的NiV毒株进行基因组测序,发现在一个局部区域和短时间内存在多个共同循环的病毒谱系,揭示了病毒演化的复杂性 [233]。甚至在1998年马来西亚的猪群疫情中,研究也发现至少存在两种不同的NiV毒株,暗示了疫情可能源于多次独立的病毒引入事件 [234]。对NiV在血液和细胞培养基中存活能力的研究则为实验室和医疗环境中的风险管理提供了关键数据,结果显示病毒在室温下密封的血液样本中异常稳定 [235]。 病毒-宿主相互作用与发病机制对NiV感染机制的理解正在从宏观走向微观。一项重要研究发现,NiV能在人血管平滑肌细胞(SMC)中高效复制,但并不引起明显的细胞病变效应(CPE),这与在内皮细胞中观察到的典型合胞体形成和细胞死亡形成鲜明对比。这一发现提示SMC可能成为病毒持续性感染的“避难所”,这或许能为部分患者出现复发性或晚发性脑炎提供一种解释 [236]。早期的体外研究也观察到,NiV在猪肾细胞、人神经元细胞、人肺成纤维细胞和人单核细胞中的复制和CPE表现各不相同,其中神经元和单核细胞在感染后几乎不向胞外释放病毒RNA,这暗示了不同细胞类型在病毒体内播散和致病中的差异化作用 [237]。 宿主因子在病毒感染周期中的作用是另一个研究热点。通过全基因组siRNA筛选,研究人员发现核仁蛋白纤维蛋白(fibrillarin)是亨尼帕病毒感染所必需的关键宿主蛋白,其缺失会严重影响病毒的RNA复制 [238]。另一项双重microRNA筛选则揭示,免疫应答相关的miR-181家族能显著促进亨尼帕病毒的进入和细胞-细胞融合 [239]。此外,有研究发现,先前感染过常见的人类副粘病毒(如麻疹病毒、副流感病毒)所诱导的T细胞,可能与致命性的NiV产生交叉反应,这表明既往的病毒感染史可能为部分人群提供一定程度的交叉保护 [240]。 通过与近缘但致病性较低的雪松病毒(Cedar virus, CedV)进行比较,可以更好地理解NiV的高致病性机制。研究发现,与NiV和亨德拉病毒(HeV)的融合蛋白(F蛋白)不同,CedV F蛋白的生物学活性同样依赖于其胞质尾部酪氨酸基序介导的内吞作用,这为解析F蛋白激活过程中的保守与特异性机制提供了线索 [241]。 新型诊断、预防与治疗策略面对NiV等高危病原体,开发快速、安全、有效的研究工具和医学对策是当务之急。基于慢病毒载体的假病毒系统,因其可在BSL-2实验室安全操作,已成为评估疫苗和药物中和活性的重要工具。研究人员不仅成功构建了表达NiV糖蛋白的假病毒系统并建立了小鼠评价模型 [242],还将此类技术应用于其他新发病毒(如SARS-CoV-2)的研究,并探索了传统草药(如高丽红参)的抗病毒效果 [243]。在诊断方面,基于探针捕获富集和高通量测序的“VirBaits”技术,能够显著提高复杂样本中包括NiV在内的多种人畜共患病病毒的检测灵敏度 [244]。 在疫苗开发领域,多种技术路线正在被探索。一项重要的疫苗设计策略是,在构建基于水泡性口炎病毒(VSV)的载体疫苗时,通过截短VSV自身的糖蛋白(如埃博拉病毒GP),可以降低其免疫原性,从而将免疫应答更集中地引向目标抗原(如NiV G蛋白),以期获得更优的保护效果 [245]。针对猪这一NiV的重要中间宿主,研究人员评估了牛疱疹病毒4型(BoHV-4)作为载体表达NiV G或F蛋白的免疫原性,发现BoHV-4载体能诱导强烈的抗体和T细胞反应,显示出作为“同一健康”策略下兽用疫苗的潜力 [246]。此外,一篇综述性文章系统回顾了NiV和HeV疫苗的研发进展,为该领域提供了全面的参考 [247]。 在治疗方面,抗体疗法是极具前景的方向。一项研究通过合理选择针对亨尼帕病毒受体结合蛋白(RBP)上不同抗原位点的人源单克隆抗体,发现这些抗体组合使用时表现出协同作用,为开发高效的抗体鸡尾酒疗法提供了依据 [248]。小分子抑制剂的开发也在进行中,研究人员基于副粘病毒F蛋白的结构相似性,筛选出能有效抑制NiV包膜蛋白介导的细胞融合的喹诺酮类衍生物,为药物设计提供了先导化合物 [249]。 结构生物学与病毒组装对病毒基本组件的结构解析是理解其功能和开发靶向药物的基础。尽管直接针对NiV,但对同属副粘病毒科的仙台病毒(SeV)的研究同样具有重要的参考价值。一项研究通过冷冻电镜技术解析了SeV“双头”核衣壳的结构,揭示了其尾对尾的组装方式、螺旋聚合机制以及一个超闭合状态的连接区域。该结构还首次直接观察到与核衣壳曲率相关的C端尾部环,为理解副粘病毒基因组的包装、复制和转录调控提供了新的结构生物学见解 [250]。这些发现补充了对NiV核衣壳和聚合酶复合物的研究,共同构成了对副粘病毒生命周期分子机制的完整图景 [251]。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号